Hydrangea petiolaris
![]()
Sponsor
Kindly sponsored by
This genus has been sponsored and new text is being prepared.
Credits
Article from Bean's Trees and Shrubs Hardy in the British Isles
Recommended citation
'Hydrangea petiolaris' from the website Trees and Shrubs Online (treesandshrubsonline.
Genus
Synonyms
- Hydrangea anomala subsp. petiolaris (Sieb. & Zucc.) McClintock
- H. scandens Maxim.
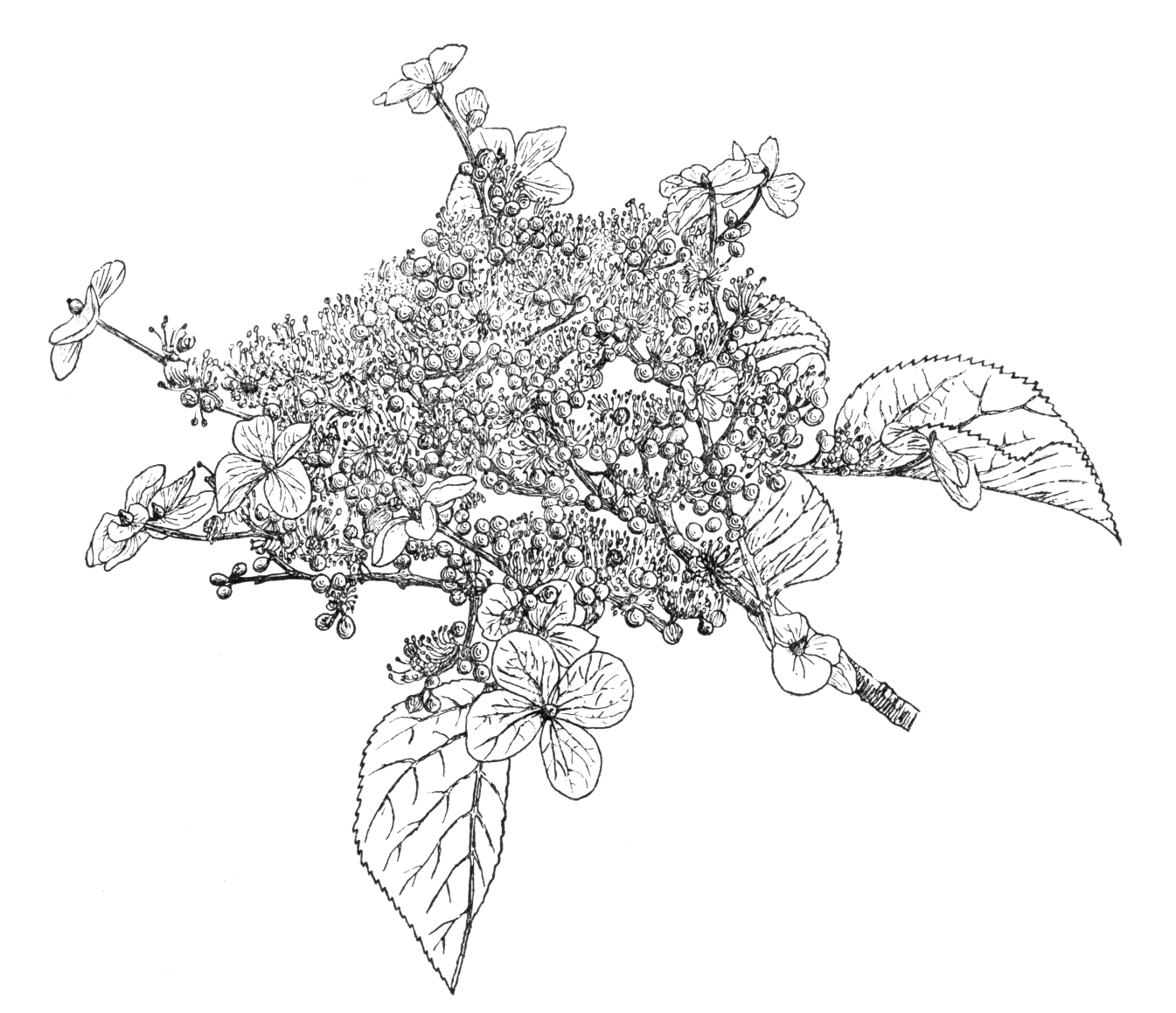
A deciduous climber, reaching in Japan to the tops of trees 60 to 80 ft high, and attaching itself closely to the trunks and limbs by means of aerial roots; young stems glabrous or hairy, older ones with peeling brown bark. Leaves roundish ovate, straight or heart-shaped at the base, and with short, tapered points; regularly, sharply, and finely toothed, 11⁄2 to 41⁄2 in. long, two-thirds to nearly as wide, dark vivid green and glabrous above, paler and often with tufts of down in the vein-axils beneath; stalk varying in length from 1⁄4 to 4 in. Corymbs expanding in June, flat, from 6 to 10 in. across, with large white sterile flowers on the margins, 1 to 13⁄4 in. across, and on stalks 1 to 11⁄2 in. long; the small fertile ones with which the centre is filled being a duller white; stamens fifteen to twenty-two; flower-stalks downy. Bot. Mag., t. 6788.
Native of Japan, Sakhalin, Quelpaert Island (Korea), and Formosa; introduced in 1878. This climber ascends trees, walls, or whatever support it has, in much the same way as ivy does. It grows vigorously, and flowers well on a wall, but a more effective way of growing it is as a bush in the open, for it is very hardy. A few plants may be put round the base of an upturned tree-stump, boulder, or even a mound, which they will soon climb over and cover. After that, the mass assumes a low, spreading, bushlike form, light and elegant in appearance, and very striking when in flower. This hydrangea is in gardens often called “Schizophragma hydrangeoides”, a name that belongs to an allied but quite distinct climber. In place of the three- to six-parted sterile blossom of the present species, the sterile flower of Schizophragma consists of a single ovate sepal, 1 to 11⁄2 in. long.
